
Par ce temps froid et venteux (l'automne se termine et l'hiver commence à pointer le bout de son nez), il est bon de penser à des lieux ensoleillés synonymes de vacances. C'est pour cela que j'aimerais vous parler de plantes associées aux paysages de cartes postales : les Palmiers.
Dans un précédent article,
j’avais évoqué l’existence de cette grande famille de plantes à fleurs assez
originale, les Palmiers, appelés aussi Arecaceae. C’est la seule famille
présente dans l’ordre des Arecales, elle-même présente dans le groupe des Commelinids
au sein des Monocotylédones (voir article précédent). Ça, c’était pour l’aspect classification et phylogénie.
Intéressons nous à présent aux particularités
de cette famille.
La famille des Palmiers regroupe
à l’heure actuelle environ 2400 espèces, réparties dans 183 genres (Dransfield
et al. 2008). Toutes ces espèces, sauf exceptions, se retrouvent au niveau de la zone tropicale à la surface de la Terre.
 |
| Carte de la répartition mondiale des espèces de Palmiers. Source : Dransfield et al. 2008 |
Il est intéressant de noter que
lorsqu’on voit un Palmier, eh bien… On le reconnait au premier coup
d’œil. N’importe quel enfant inscrit à l’école primaire dessinera, si on
lui demande de représenter une île déserte, quelque chose d’approchant :
 |
| Trois vaguelettes, un soleil, un monticule de sable... et un Palmier : c'est la représentation classique d'un île déserte [source] |
Et du même coup, on se rend
compte que l’imagerie du Palmier est
connue depuis notre plus tendre enfance. Mais si c’est une chose de représenter
un Palmier en trois coups de crayon, c’en est une autre de savoir ce qui se
cache derrière ces feuilles vertes qui protègent le naufragé des rayons du
soleil.
Savez vous en effet que les Palmiers ne sont pas des arbres ?
Eh oui, ils ne possèdent pas de tronc à proprement parler ! Mais alors,
comment est-il possible que ces plantes mesurent parfois plusieurs mètres de
haut et soit dures comme du bois… sans en être réellement ?
Reprenons les choses depuis le
début. La croissance des plantes est initiée (dans un premier temps) aux
extrémités de l’individu. En particulier, on appelle les zones de croissance des méristèmes. On trouve ces méristèmes au niveau des tiges (méristèmes caulinaires) et au niveau
des racines (méristèmes racinaires).
Ces zones sont ce que l’on appelle communément les bourgeons.
 |
| Schéma d'une plante avec la localisation des méristèmes. Modifié d'après [source] |
Oui mais voilà, si on s’arrêtait
là, tout serait bien trop simple. Certaines plantes possèdent deux types de méristèmes : primaires et secondaires. Tandis que
les méristèmes primaires permettent la croissance
en longueur de la plante et sont présents au tout début de sa croissance, à
l’intérieur même de la graine, les méristèmes secondaires se mettent en place…
secondairement, et permettent la croissance
en épaisseur de la plante. Ces méristèmes sont appelés cambium (ou Assise Génératrice Libéro-Ligneuse) et phellogène (ou Assise Génératrice Subéro-Phellodermique).
 |
| Schéma d'une coupe dans un tronc, montrant les différentes couches produites par le cambium et le phellogène. Les flèches verticales montrent les zones de croissance cellulaire en épaisseur. [Source] |
D’après la figure précédente, on
peut voir que le tronc d’un arbre est constitué de plusieurs couches successives. Si on regarde en
commençant par la partie visible de l’arbre, à l’extérieur, la première couche
rencontrée est l’écorce. Cette
couche imperméable et protectrice est constituée par le liège, ou suber, qui est produit par le phellogène (ou AGSP, voir plus haut) qui est donc le premier
méristème secondaire rencontré chez un arbre, en partant de l’extérieur. A titre
d’information, les bouchons de bouteille sont faits en liège, qui est en
réalité l’écorce des Chênes Quercus suber,
récoltée annuellement. Ce matériau est utilisé pour cet usage du fait de sa grande imperméabilité.
Si on avance vers l’intérieur du
tronc, on tombe sur le phelloderme,
une mince couche cellulaire produite elle aussi par le phellogène. Cette couche
fait partie de l’écorce mais reste
généralement sans rôle particulier.
La couche suivante est le liber, aussi appelé phloème secondaire : c’est un
tissus qui conduit la sève élaborée,
riche en sucres, depuis les feuilles vers les racines. Il est produit par le cambium (ou AGLL, voir plus haut), le
second méristème secondaire, vers l’extérieur de la plante.
La couche suivante est l’aubier : c’est la partie vivante
du bois. Il s’agit du xylème secondaire,
également produit par le cambium vers l’intérieur de la plante. Il sert à
conduire la sève brute, constituée
essentiellement d’eau et de sels minéraux, depuis les racines jusqu’aux
feuilles. Par la suite, les cellules de l’aubier vont se remplir de lignine et
subir ce que l’on nomme la duraminisation :
on obtiendra ainsi le bois de cœur, encore appelé duramen.
 |
| Coupe transversale schématique d'un tronc. Notez qu'ici, l'écorce rassemble le suber, le phellogène et le phelloderme. [Source] |
La production de bois permet, à
postériori, la formation d’un tronc,
suite à la rigidification des tissus végétaux. Evolutivement, la présence du
tronc peut s’expliquer par une constante compétition par les plantes au niveau
de l’occupation du volume par les
structures photosynthétiques : la présence d’un
« squelette » constitué par le tronc permet à la plante de déployer
ses feuilles dans toutes les directions et ainsi de maximiser leur exposition à
la lumière. Vous imaginez bien qu’avoir un étagement
des feuilles permet d’augmenter la surface
photosynthétique pour une même surface au sol… C’est un peu comme avec nos
immeubles : on garde la même surface au sol qu’une maison d’un étage, mais
on empile des appartements, donc tout le monde au final a plus de place pour
vivre.
L’augmentation de la taille à
l’aide d’un tronc permet également d’accroitre la capacité de dispersion des
fruits et des graines. C’est logique : lancez une balle depuis le pied
d’un arbre et depuis sa cime et vous verrez qu’elle parcourra plus de distance
si vous vous situez au sommet de l’arbre.
Tout ça c’est bien joli, mais… les Palmiers n’ont pas de tronc à
proprement parler. En effet, une des particularités des Monocotylédones (voir
article précédent) est d’avoir perdu
secondairement, au cours de l’évolution, la capacité de former des structures secondaires. Ces plantes ne
possèdent tout simplement pas de méristème secondaire ! Comment est-il
possible, dans ce cas, que les Palmiers soient des plantes dont la taille
dépasse aisément plusieurs mètres ?
Eh bien, ces plantes possèdent d’autres structures qui leurs
permettent aussi de se hisser vers la lumière. La partie analogue à un tronc
d’arbre chez un palmier s’appelle un stipe,
et c’est ce stipe qui permet d’étendre la couronne de feuille au soleil, loin
au dessus du sol.
Ce stipe est constitué d’un grand nombre de fibres,
rigidifiées par la lignine (toujours elle). Dans un stipe, on trouve un très
grand nombre de fibres conductrices
de sève, associées souvent avec un ensemble de fibres de soutien.
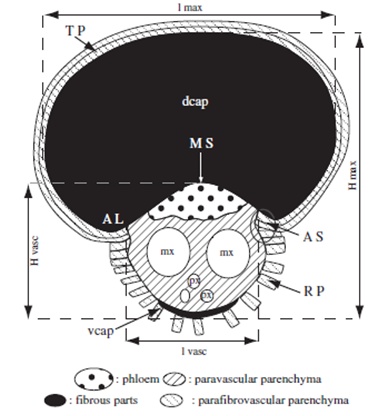 |
| Détail d'une fibre conductrice associée à une fibre de soutien (en coupe transversale). En noir, les fibres de soutien ; en pointillés, le phloème ; en blanc (mx et px), du xylème ; en hachuré, du parenchyme (c'est un tissus de soutien). Il n'y a pas de tissus secondaires. Source : Thomas & De Franceschi 2012 |
 |
| Voici les fibres, replacées dans le contexte entier du stipe (à g., extérieur, à dr., intérieur), pour les deux types d'organisation chez les Palmiers (Thomas & De Franceschi 2012) |
Bien qu’il n’existe pas de croissance secondaire à
proprement parler chez les Palmiers, l’accumulation et l’agrégation des fibres
enchevêtrées entre elles permet au stipe d’augmenter petit à petit en épaisseur
(surtout au niveau de la base), ce qui confère au Palmier dans son entier sa
stabilité et sa solidité.
D’un point de vue de la
croissance, comme je l’ai dit plus haut, les Palmiers n’ont pas de méristèmes secondaires donc pas de croissance secondaire
à proprement parler : un seul méristème initie la croissance de la plante,
d’où la présence d’une couronne de
feuille unique au sommet et l’absence de branches. Chaque année, les
feuilles de l’année précédentes meurent et le Palmier initie la croissance de nouvelles feuilles juste au dessus des
anciennes. On pourrait dire ainsi qu’un Palmier est le résultat d’un emboitement successif de feuilles,
année après année. Mais on ne peut pas parler de vrai tronc car il n’y a pas
d’accroissement en largeur, suite à l’absence de méristèmes secondaires.
 |
| Différentes coupes transversales de stipes de Palmiers, mettant en évidence les fibres vasculaires. [Source] |
Mais il ne faut pas croire que
les Palmiers sont tous pareils : c’est une famille de plantes à fleurs où
l’on trouve une diversité de formes,
de tailles, de couleurs impressionnantes. Ainsi, si la plupart des Palmiers
possèdent des stipes « conventionnels », on trouve des Palmiers rampants et grimpants. Si
certains Palmiers peuvent atteindre 15 mètres , d’autres sont nains et ne dépasseront jamais un mètre de hauteur.
 |
| Calamus nabariensis, détail du stipe (à g.) et Calamus longipina, vue d'ensemble (à dr.) : deux Palmiers grimpants. [Source] |
 |
| Chamaerops humilis (à g.) et Actinorhytis calapparia (à dr.) : notez la différence de taille... [Source] |
Certains Palmiers ont des stipes à épines alors que d’autres
possèdent un stipe complètement lisse ou bien fibreux.
 |
| Calamus wailong (à g.) et Howea forsteriana (à dr.). Notez les stipes épineux à divers degrés du Calamus et celui parfaitement lisse du Howea. [Source] |
Mais le plus impressionnant à mon
sens reste le Coco-Fesse (nom commun du Lodoicea
maldivica), qui possède un fruit en forme de… bin, je vais pas vous faire
un dessin, qui peut peser jusqu’à 25
kilos !
 |
| Fruit de Lodoicea maldivica, avant (à g.) et pendant (à dr.) la germination. [Source] |
Les fruits du Cocotier Cocos nucifera (les noix de Cocos) sont
également capables de prouesses au
niveau de la capacité de germination : on les retrouve parfois à des
milliers de kilomètres de distance de leur point de départ, portés par l’océan.
S’ils échouent sur une plage tropicale, ils sont capables de germer très
rapidement et de prendre racine sur le rivage.
 |
| Une noix de Coco germe après avoir été transportée par la mer. [Source] |
Chez les Palmiers, on trouve
aussi une grande diversité de feuilles, bien qu’elles soient toutes construites
sur le même modèle de base : lors de leur formation, elles sont repliées
comme un pliage d’origami, puis elles se déplient pour prendre leur forme
définitive. On distingue souvent deux grands types de feuilles, parmi lesquels
on trouve toutes sortes de variations morphologiques : il s’agit des
feuilles pennées (comme des plumes) ou palmées (comme une main).
 |
| Areca vestiaria (à g.) possède des feuilles pennées, tandis que Serenoa repens (à dr.) possède des feuilles palmées. [Source] |
Pour conclure, je dirais qu’au
sein du règne végétal, il ne faut pas se fier aux apparences… et que même s’ils
en ont l’air, les Palmiers ne sont pas
des arbres et recèlent une diversité
insoupçonnée au premier regard.
Bibliographie
Dransfield et al. 2008. Genera Palmarum. Royal Botanic Garden, Kew
Thomas R & De Franceschi D. 2012. First evidence of fossil Cryosophileae (Arecaceae) outside the Americas (early Oligocene and late Miocene of France): Anatomy, palaeobiogeography and evolutionary implications. Review of Palaeobotany and Palynology. 171:27-39
Pour les photo, l'immense ressource d'Internet en général et le site http://palmweb.org/ en particulier.







{kind=link}