
Certains voient peut-être la zoologie comme une vieille discipline qui a fait son temps. L’étude de la morphologie elle aussi est vue parfois comme une ancienne discipline. La majorité des observations sur les animaux auraient été faites. C'est tout à fait l'inverse, beaucoup de mystères subsistent et beaucoup de choses sont découvertes encore comme de nouveaux groupes d'animaux inconnus. Mais une chose passionnante est l'impact de la phylogénie. En effet, l'interprétation de la morphologie et des relations évolutives entre animaux change totalement selon l'arbre des animaux qu'on prend en compte. Je vais essayer de vous présenter brièvement quelques grands changements dans cette aventure intellectuelle...
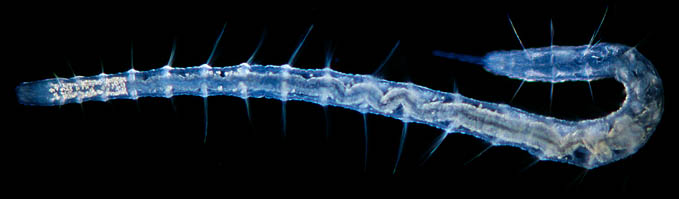
Parlons déjà du mystère des Siboglinidae. Vous avez peut-être entendu parler des vers Vestimentifères, ce sont des Siboglinidae. Les vestimentifères sont de grands vers des abysses avec une amusante touffe rouge à l’avant et qui vivent en symbiose avec des bactéries sulfuriques. Pendant longtemps on a rapproché ces animaux des échinodermes (les échinodermes comprennent entre autre les étoiles de mer et oursins) et des entéropneustes (comprenant le balanoglosse ou « ver-pénis » voir l'article sur les animaux obscènes). Cela était bien argumenté grâce à la structure du système nerveux, la position des organes reproducteurs et le développement. Autant dire que les arguments envoyaient du pâté. Mais en 1964 une drôle d'observation vint mettre le fouillis là dedans : jusqu'alors la partie postérieur (l’arrière, le cul quoi) de ces vers n’avait jamais été observée. Et c'était quasiment sans appel, cette partie postérieure était clairement celle d'une annélide. Les annélides vous les connaissez, le ver de terre en est une. Cette partie postérieure portait des « poils » (ou soies) et était constituée d’anneaux comme ce qui est communément le cas chez les annélides. Or cette organisation ne se retrouve pas sur le reste du corps des Siboglinidae. Mais les zoologistes sont de petits têtus et certains négligèrent cette observation. Plus tard elle fut progressivement mieux acceptée et maintenant les zoologistes en mettraient leur main à couper, les Siboglinidae sont des annélides ! La ré-interprétation de la morphologie mais aussi les arbres obtenus grâce à l'ADN coïncident finalement. Réinterprétation de la morphologie ? Mais elle n'indiquait pas ça au départ pourtant ! Certains zoologistes ont ré-étudié avec plus d'attention les même caractères pour se rendre compte que... Par exemple le système nerveux qui était supposé dorsal (comme chez nous et les entéropneustes, les vers-pénis) est en fait ventral ! Un beau retournement de cerveau pour un beau retournement de situation ! Notez qu'au sein des animaux à deux côtés (bilatériens) avec un avant et un arrière, les Siboglinidae ont fait un voyage considérable d'un bout à l'autre de l'arbre. Toutes ces histoires pour un bout de cul... Finalement les histoires de phylogénie ne sont pas si différentes des histoires d'humains : on en revient toujours en dessous de la ceinture !
 |
| Sibolinidae et annélides : en haut à gauche, des vers vestimentifères (source de l’image : vers vestimentifères). En haut à droite un dessin de l’opisthosome perdu des Siboglinidae… Leur cul quoi pour faire moins classe (source de l’image : Pleijel et al. 2009). En bas une photo d’une annélide. Remarquez la structure répétée et la présence de « soies ». (Source de l’image : joli clitellate) |
 |
| A gauche une image de Xenoturbella (source de l’image : Xenoturbella). A droite une étoile de mer (source de l’image : étoile de mer). Oui, les deux ne se ressemblent pas (remarquez qu’en fait les deux ne ressemblent pas à grand chose en général) mais ils sont plus proches entre eux qu’ils ne le sont des vertébrés. Et nous les vertébrés sommes plus proches d’eux que nous le sommes des poulpes ou des insectes ! |
Les myxozoaires ont été eux considérés pendant un certain temps comme des "protozoaires" c'est à dire des êtres unicellulaires. En tout cas des organismes qui ne sont ni des animaux, ni des plantes, ni des champignons, ni des bactéries (sympa comme définition hein ?). Pourtant ces organismes ont plusieurs cellules. Pourquoi cette injustice ? A cause du mode de reproduction par spores inhabituel chez les animaux et d’une structure très simple. Ces organismes parasites de vertébrés aquatiques (ben sur ce blog le mot "poisson" est interdit ! ) et d'autres organismes ont deux cellules avec un harpon au bout d'un lasso qu'elles peuvent dévaginer (éjecter) pour ralentir leur chute dans le tube digestif de l'organisme qu'ils parasitent... Peut-être que certains amateurs de zoologie passant par là auront pensé à un certain groupe d’animaux... D'autres organismes ont des cellules qui ont un lasso à harpon, les cnidaires. Vous en avez fait probablement la piquante expérience, les méduses étant des cnidaires. Les myxozoaires se sont donc retrouvés classés chez les cnidaires, ce sont finalement des animaux proches des méduses. Mieux même, on saurait les placer précisément au sein des cnidaires. Et ils seraient d'ailleurs proches d'autres cnidaires parasites. Cependant le cheminement n'a pas été aussi simple. Si on a longtemps hésité à les considérer comme des animaux, les premières analyses utilisant l'ADN les ont très vite identifié comme des animaux mais plus proches des bilatériens que des cnidaires malgré les cellules à harpons (comme, on la pensé pour les xenoturbellides, la proximité des bilatériens est une place convoitée…). La découverte d'un myxozoaire à la forme d'un ver vint apporter de l'eau au moulin de cette hypothèse, en effet, fondamentalement, les animaux bilatériens avec un avant et un arrière sont des vers (oui, vous lecteurs, vous êtes des vers !). D'autres analyses de l'ADN vinrent ensuite réfuter cette hypothèse et revinrent à l'idée que les myxozoaires sont bien des cnidaires parasites. Imaginez cependant : un organisme qui n'était au départ pas considéré comme un animal se retrouve finalement bien identifié comme appartenant à un sous groupe d'animal et même à un sous groupe de ce sous groupe d'animal. Ca en fait un voyage conceptuel !
 |
| En haut à gauche des myxozoaires… Dur de penser à des méduses très modifiées ! (source de l’image : myxozoaires qui r’semblent à rien). En bas à gauche le myxozoaire en forme de ver : Buddenbrockia plumatellae ! Celui qui a mené les zoologistes sur une fausse route (source de l’image : Buddenbrockia plumatellae le fourbe). A droite, une méduse du genre Clytia, probablement plus proche des myxozoaires que des méduses que vous connaissez et qui vous piquent sur la plage… Pourtant elle leur ressemble bien plus (source de l’image : Clytia la tite méduse). |
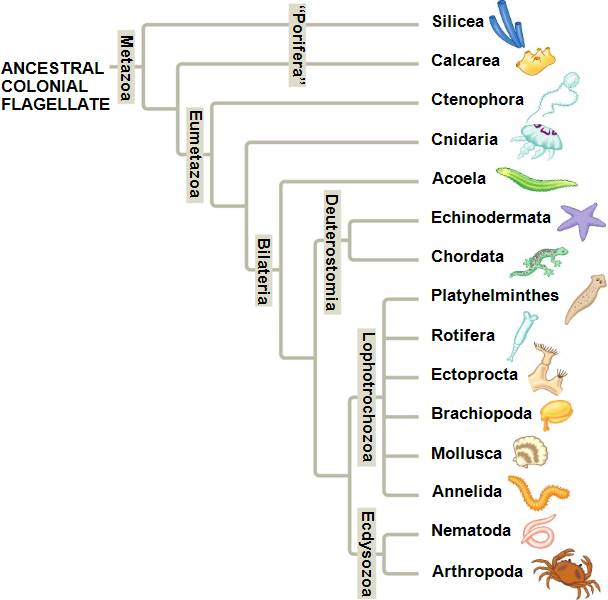
Jusque là je vous ai parlé de sous groupes d'animaux qui se baladaient, comprenant pour ceux-là quelques espèces chacun. Mais il y a des remaniements bien plus importants. Souvenez-vous des annélides dont je parlais dans la première partie et dans cet article sur les organismes obscènes (Vous pouvez aussi vous référer à cet article ici). Ces animaux sont segmentés (plus exactement métamérisés), c'est à dire qu'ils sont constitués d'unités morphologiques distinctes mais similaires mises à la suite les unes des autres. Imaginez en fait une chaîne. D'autres organismes sont du même genre : les arthropodes comprenant les insectes, arachnides et les « crustacés » (qui comprennent tout ce que vous appelez crustacés mais qui n'existent pas plus que les poissons puisqu’ils devraient comprendre les insectes !). On peut les imaginer aussi comme une chaîne ou un ensemble d'anneaux similaires. Bref ce joyeux groupe des arthropodes est organisé selon la même structure que les annélides. Quoi de plus normal que de les regrouper dans un groupe appelé "Articulata". Petit bémol, les mollusques ont un développement très proche de celui des annélides. Rhoo on va dire que ce n'est pas trop grave. Mais l'analyse de l'ADN sépara nos deux conjoints. L'ADN seulement ? Une analyse morphologique en 1992 le fit aussi plusieurs années avant l’étude de l'ADN et réfutant l'hypothèse "Articulata" montrant ainsi que la métamérie était apparue indépendamment dans ces deux groupes, que c’est une convergence. Mais cette étude ne fut pas beaucoup reprise, les auteurs n'ayant probablement pas réalisé l'impact de leur découverte. Régulièrement grâce à l'ADN, on remarqua alors deux grand groupes : les Ecdysozoa regroupant les animaux qui muent dont les arthropodes comme les insectes ou d'autres étranges vers comme les nématodes ou les priapuliens. Un autre groupe fut mis en évidence, celui des lophotrochozoaires, dont on ne trouve au final pas vraiment de bon caractère morphologique pour les décrire dans leur ensemble. Les lophotrochozoaires sont un groupe comprenant des animaux tous plus étranges les un que les autres : annélides déjà, mollusques, platheminthes mais aussi rotifères, gnatostomulides, gastrotriches, phoronidiens et autres étranges joyeusetés animales. Si la classification au sein des ecdysozoaires, bien qu'encore en discussion, ne semble pas être trop bordélique, ce n'est pas le cas chez les lophotrochozoaires où les relations de parentés sont aussi confuses que la prononciation de ce mot et de nombreuses découvertes restent encore à faire.
 |
| L’arbre des animaux très simplifié (et encore en discussions hein). Annélides et arthropodes sont entourés en rouge. Remarquez qu’ils sont éloignés. Et oui, à force d’être trop attachés à un moment il faut prendre ces distances ! (Source de l’image : arbre des animaux un peu beaucoup simplifié) |
Pour aller plus loin :
Tous les ouvrages de zoologie générale devraient faire l'affaire ! :
Classification phylogénétique du vivant de Guillaume Lecointre et Hervé le Guyader, 2006, éditions Belin.
Invertebrates, second edition de Richard C. Brusca et Gary J. Brusca , 2003, éditions Sinauer.
Les invertébrés marins méconnus de Jean Loup d'Hondt, 1999, éditions Institut océanographique
Précis de siences biologiques, zoologie, I Invertébrés de P. P. Grassé, R. Poisson et O. Tuzet, 1961, éditions Masson et Cie.
Quelques articles :
-Bourlat S. J., Nielsen C., Lockyer A. E., LittlewoodD. T. J. et Telford M. 2003. Xenoturbella is a deuterostome that eats molluscs. Nature, 424, 925-928.
-Eernisse D.J., Albert J.S. et Anderson F.E. 1992. Annelida and arthropoda are not sister taxa: a phylogenetic analysis of spiralian metazoan morphology. Systematic biology, 41(3), 305-330.
-Mallatt J., Waggoner Craig C. et Yoder M.J. 2010. Nearly complete rRNA genes assembled from across the metazoan animals: Effects of more taxa, a structure-based alignment, and paired-sites evolutionary models on phylogeny reconstruction. Molecular Phylogenetics and Evolution, 55 : 1-17… (Ben ça c’est du nom d’article à rallonge ! )
-Manuel M. 2009. Evolution animale : les péripéties de la phylogénie. Encyclopédie Universalis, 134-149.
-Pleijel F., Dahlgren T.G. et Rouse G.W. 2009. Progress in systematics: from Siboglinidae to Pogonophora and Vestimentifera and back to Siboglinidae. C. R. Biologies, 332, 140-148.
 |
Au final un arbre des animaux bien moins simplifié publié en 2010… Comme quoi ben c’est pas facile de s’y retrouver au milieu de tout ça ! (Source de l’image : Mallat et al. 2010)














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}